Thomas J. Givnish
Henry Allan Gleason Professor of Botany and Environmental Studies
Ph.D. (1976) Princeton University • 315 Birge Hall • 608-262-5718 • givnish@wisc.edu
Plant ecology and evolution; adaptive radiation and molecular systematics;
phylogeography; physiological ecology; landscape dynamics

Physiological ecology and biomechanics are major areas of interest.
I am currently coordinating efforts with Kate McCulloh, Mark Adams, Tom Buckley, Duncan Smith, and Amanda Salvi to study the adaptations of 88 traits in 10 Eucalyptus species to different portions of a steep rainfall gradient in Victoria, Australia, using a series of four common gardens along that gradient. We are testing a large series of hypotheses regarding trait-environment and trait-trait relationships based on economic theory, and assessing the integration of photosynthetic, hydraulic, biomechanical, and allocational traits. We use P/Ep – the ratio of precipitation to pan evaporation – as an index of relative moisture supply (the ratio of input to possible demand), to quantify the climatically determined moisture supply characteristic of individual common gardens and the range of individual species. We are also asking whether these species show adaptive crossover in realized height growth, with species having an advantage only in parts of the gradient they dominate in nature.
In 2014, working with another set of colleagues, I showed that maximum tree height on this same Victorian climatic gradient increased more or less linearly with P/Ep.
Amanda Salvi recently completed a ground-breaking study showing that mesophyll photosynthetic sensitivity (MPS) – the decline in photosynthetic capacity with drops in leaf water potential, after the effects of shifts in stomatal conductance have been removed – decreases in regular fashion as the relative moisture supply across a species’ range declines, but at the cost of reduced photosynthetic capacity at full hydration. That is, various measures of MPS and maximum photosynthetic capacity increase with species P/Ep. Furthermore, Salvi et al. have found that species with greater MPS (i.e., those from moister habitats) are more isohydric; that is, they control their stomatal conductance more rigorously. These results are consistent with my model for optimal stomatal conductance but not the widely cited ∂A/∂E model, and have profound implications for plant adaptation, resource allocation, and growth, for the relationship of plant traits to climate, and – ultimately – for global patterns of climate itself. We will be pursuing these important results both theoretically, in terms of new optimality models, and empirically in other plant groups.
Our earlier work on Eucalyptus included a new conceptual framework for constraints on maximum tree height. Data from the Victorian rainfall gradient in showed a tight relationship between maximum height of Eucalyptus species and relative moisture supply as measured by P/Ep. Variation in the stable carbon isotope ratios of trees along this gradient implicate both hydraulic constraints and biomechanical limitations as setting maximum tree height.
Jared Beck and I recently published study showing the differential spatial distribution of three forest herbs within a deciduous forest relative to soil depth and tree boles, consistent with theories based on differences in leaf height, rooting depth, and leaf photosynthetic capacity.
This year I re-assessed Henry Horn’s classic The Adaptive Geometry of Trees, pointing out a previously unrecognized flaw in its core model, and suggesting alternative explanations to account for patterns in the number of leaf layers held by early vs. late successional tree species. I am eager to recruit a study or post-doc to test some of these new models.
Robert Wernerehl and I continue to explore the distribution of prairie species in relation to soil moisture, nutrient supplies, and (especially) soil mechanical impedance, and to use drought and flooding experiments to assess the causes of the differential distribution of dominant prairie grasses along moisture gradients.
Rebecca Montgomery and I are studying the ecology and evolution of photosynthetic light adaptations in the Hawaiian lobeliads (see Adaptive Radiation). These investigations recently included collaborative work with Christine Scoffoni and Lawren Sack on adaptation to light x moisture availability. Rebecca, Omar and Krista Lopez, and I are analyzing how time of leafing and hydraulic conductance influence shade tolerance in trees of the southern Appalachians. Early leafers obtain a “spring subsidy” of carbon by photosynthesizing a few extra days or weeks under an open canopy, which may allow them to persist in microsites that are shadier in midsummer. To do so, however, they must have narrow xylem elements to weather late frosts. Such elements resist cavitation but are hydraulically inefficient. Late leafers with more efficient vessels may be able to sustain higher rates of transpiration (and hence photosynthesis) in sunnier microsites, even if they have higher whole-plant compensation points. We have published some tests of these hypotheses and are developing others, based on studies in Great Smoky Mountains National Park and Nantahala National Forest.
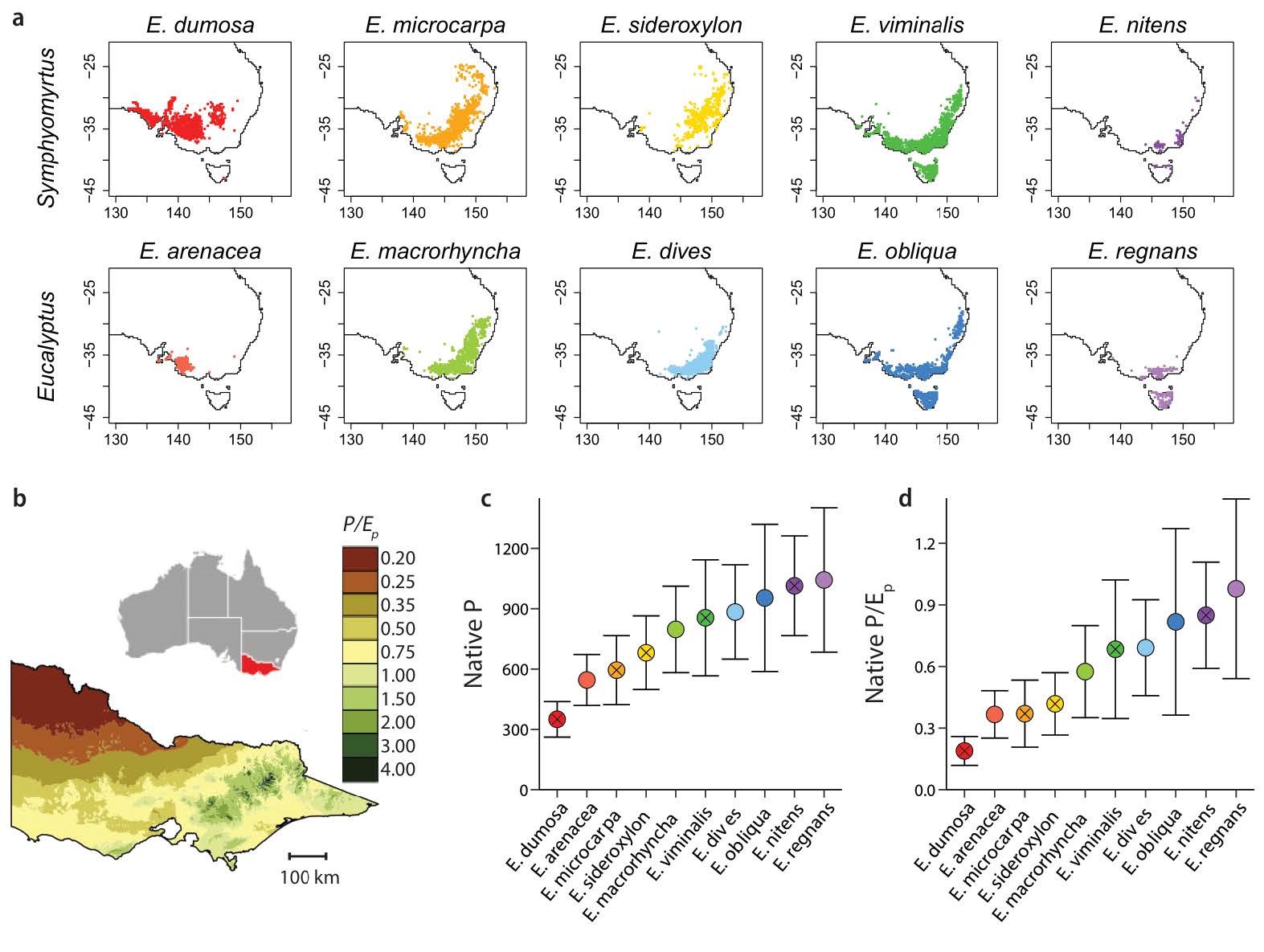
Photographs: TOP - Portrait of the scientist as a young man among Nymphaea; Cyanea floribunda (Lobeliaceae), a highly shade-adapted lobeliad from windward Hawai`i, in wet forests of the Ola`a Tract of Hawaii Volcanoes National Park; Pinus lambertiana (Pinaceae), a common associate of the giant redwood in the Sierra Nevada and among the tallest trees on earth; shade-tolerant saplings of Ostrya virginica leafing out under the as-yet-unopened canopies of Quercus rubra and other late-leafing, shade-intolerant trees at Clifty Falls Park, Indiana; Eucalyptus diversicolor (karri) temperate rain forest in southwestern Australia, dominated by the second tallest species of flowering plants in the world. BOTTOM - Native distributions and climate data of the 10 Eucalyptus species (after Salvi et al. 2021). (a) Geographic distributions across Australia, ranked by subgenus (rows) and native P/Ep (least = driest on the left). (b) Map of Victoria, from Givnish et al. 2014, showing contours of relative moisture supply (P/Ep). (c) Mean ± s.d P/Ep and (d) mean annual precipitation across each species range. Crossed circles = subgenus Symphyomyrtus.
© 2021 University of Wisconsin Department of Botany
Last updated: 27 November 2021



